ISBN 978-85-85905-15-6
Área
Bioquímica e Biotecnologia
Autores
Sousa Oliveira, M. (UNIVERSIDADE ESTADUAL DE GOIÁS) ; Araújo Toledo, J. (UNIVERSIDADE ESTADUAL DE GOIÁS) ; Júnio Cantuária de Oliveira, A. (UNIVERSIDADE ESTADUAL DE GOIÁS) ; Cristina Garcia Martin Didonet, C. (UNIVERSIDADE ESTADUAL DE GOIÁS) ; Lázaro Faleiro Naves, P. (UNIVERSIDADE ESTADUAL DE GOIÁS)
Resumo
O uso de fertilizantes químicos pode impactar negativamente o meio ambiente. Dessa forma bactérias promotoras de crescimento vegetal vêm se destacando por sintetizar compostos que auxiliam no crescimento e produtividade de diversas culturas. Entre esses compostos está o Ácido Indol-3-Acético (AIA). Este trabalho objetivou quantificar a produção de AIA por 20 bactérias da família Rhizobiaceae isoladas de nódulos de raízes de plantas de arroz Sequeiro (Oryza sativa) quando cultivadas em meio batata enriquecido com triptofano, através da técnica colorimétrica com reagente de Salkowski. Todas as bactérias produziram AIA com quantidades variando entre 0,207 a 33,212 μg.mL-1, sendo que cinco foram identificadas como fortes, onze moderadas e quatro de baixa produção.
Palavras chaves
Ácido Indol-3-Acético; Família Rhizobiaceae; Arroz Sequeiro
Introdução
Melhorar a fertilidade do solo e a produtividade das culturas reduzindo os impactos negativos dos fertilizantes químicos no ambiente é imprescindível para a agricultura. Rizobactérias promotoras de crescimento vegetal e bactérias simbióticas fixadoras de nitrogênio são uma alternativa que visa atender a essas necessidades (BABALOLA, 2010). Uma alternativa química para regular o crescimento de plantas e consequentemente conseguir alta produção agrícola é através do uso de fitormônios. Dentre esses compostos destacam se as auximas, citocininas e as giberilinas (NETO et al, 2004). Estes compostos são aplicados de forma exógena durante o cultivo de diferentes culturas. As concentrações utilizadas destes fitorreguladores exógenos variam entre 1,0 e 10,0 μM, concentrações elevadas se comparadas às concentrações dos fitormônios endógenos (CHEN et al, 2010). Uma alternativa ao uso de fitormônios sintéticos é a utilização de bactérias promotoras de crescimento vegetal, as quais possuem a capacidade de produzir hormônios, vitaminas e fatores de crescimento que auxiliam no desenvolvimento de diferentes tipos de plantas (DIMKPA et al, 2012). Essas bactérias exercem efeitos positivos diretos e indiretos no desenvolvimento das plantas. Diretamente se observa o aumento da solubilização e entrada de nutrientes e a produção de reguladores de crescimento vegetal. Indiretamente se observa a supressão de patógenos devido à produção de antibióticos (KUSS et al, 2007). As auxinas destacam-se dentro do grupo de reguladores de crescimento. Esses compostos são um tipo de hormônio vegetal produzidos pelas plantas e que são utilizados na transdução de sinal intracelular e intercelular. As auxinas estão envolvidas em processos de desenvolvimento, incluindo divisão e diferenciação celular, fototropismo, dominância apical e diferenciação vascular (SUGAWARA et al, 2009). A presença de auxinas provocam alterações morfológicas, tais como, o aumento do comprimento da raiz, o número de pêlos radiculares e raízes laterais. Esses compostos estimulam na planta respostas rápidas, como o aumento do alongamento celular e respostas lentas entre elas a divisão e diferenciação celular (BONILLA, 2011). O ácido indol-3-acético (AIA) é a auxina natural mais abundante e estudada atualmente. Ela atua como uma molécula de sinalização e é um regulador chave, exercendo inúmeros papeis no desenvolvimento da planta (SUGAWARA et al, 2015). A produção de AIA por bactérias vem sendo relatada em diversos estudos (PRINSEN et al, 1991), (KUSS et al, 2007), (TSAVKELOVA et al, 2012). Existem duas vias de biossíntese propostas para AIA, uma via dependente de triptofano e outra independente dele. Na via dependente de triptofano diferentes caminhos vem sendo postuladas, entre eles estão a via indol-3 acetamida (IAM), via do ácido indol-3-pirúvico (AIP), via triptamina (TAM), e a via indol-3-acetaldoxima (IAOx) sendo as duas primeiras as mais observadas (MANO, NEMOTO, 2012), (BARTEL, 1997), (DUCA et al, 2014). Na via indol-3-acetamida (IAM) ocorre uma reação de dois passos, onde o triptofano é inicialmente convertido a IAM pela enzima triptofano monooxigenase. Posteriormente há a conversão do IAM para AIA em uma reação catalisada pela enzima indol-3-acetamida hidrolase (BARTEL, 1997). Na via do ácido indol-3-pirúvico (AIP) inicialmente, o triptofano é desaminado a AIP por uma aminotransferase. Subsequentemente, uma enzima descarboxilase converte em AIP a indol-3-acetaldeido, o qual é então oxidado a IAA por aldeído desidrogenase, ou enzimas oxidase (DUCA et al, 2014). A quantidade de AIA produzida pelas bactérias é influenciada por diferentes fatores, entre eles a localização dos genes responsáveis pela biossíntese (BRANDLAND, LINDOW, 1996). Fatores abióticos como pH, estresse osmótico, variações na concentração de oxigênio, temperatura e limitação de carbono também influenciam na produção de AIA (SPAEPEN, VANDERLEYDEN, REMANS, 2007). A produção de fitormônios por rizóbios durante o processo de nodulação vem sendo relatada como uma possível forma de estímulo ao crescimento vegetal (STROSCHEIN et al, 2011). PRINSEN e colaboradores (1991) observaram que na presença de flavonoides normalmente secretados pelas plantas durante o processo de nodulação ativam não somente a expressão de genes de nodulação (Nod) na bactéria mas também causam um significativo aumento na produção de AIA. (PRINSEN et al, 1991). Associadas a estas informações este trabalho teve como objetivo avaliar a produção de AIA por bactérias da família Rhizobiaceae isoladas a partir de nódulos simbióticos presentes em raízes de arroz sequeiro (Oryza sativa) cultivado no cerrado goiano.
Material e métodos
Para a condução do experimento foram selecionados 15 isolados de plantas de arroz cultivadas no cerrado goiano, obtidas a partir do isolamento em meio específico (YM) para bactérias da família Rhizobiaceae. Os microrganismos estão catalogados e estocados na coleção do Laboratório de Bioquímica da Universidade Estadual de Goiás. Foram selecionados os isolados ISR5, ISR25, ISR30, ISR31, ISR40, ISR54, ISR58, ISR59, ISR65, ISR73, ISR80, ISR82, ISR127, ISR132 e ISR134. Também foram utilizadas cinco estirpes controle obtidas junto a Embrapa Arroz e Feijão de Santo Antônio de Goiás. Foram usadas 4 bactérias do gênero Rhizobium BR322 (Rhizobium tropici) , BR520 (Rhizobium tropici), BR10055 (Rhizobium galegae), CFN42 (Rhizobium etli), 1 do gênero Mesorhizobium BR522 (Mesorhizobium mediterraneum). O meio de cultura utilizado no experimento foi o meio Batata, composto por caldo cozido de 200 g de batata inglesa, 2,5 g.L-1 de ácido málico, 2,5 g.L-1 de açúcar cristal comercial, 0,000001 g.L-1 de biotina, 2 mL de solução de micronutrientes (0,2 g.L-1 de molibidato de sódio, 0,235 g.L-1 de sulfato de manganês, 0,28 g.L-1 de ácido bórico, 0,008 g.L-1 de sulfato de cobre e 0,024 g.L-1 de sulfato de zinco) com pH 6,8. (DOBEREINER, ANDRADE, BALDANI, 1999). O meio de cultura foi suplementado com L-triptofano na concentração de 0,25 g.L-1 como precursor para a síntese de AIA (DUCA et al, 2014). Para quantificar as auxinas produzidas pelas bactérias selecionadas, alíquotas de 500 µL de inóculo de cada um dos isolados bacterianos, ajustadas ao correspondente 0,5 da escala de McFarland, foram colocadas para crescimento em 20 mL de meio batata suplementado com triptofano. Os recipientes foram incubados a 30 °C, 140 RPM por 62 horas. Após o período de crescimento cada uma das culturas homogeneizadas foram transferidas para tubos e centrifugadas a 10.000 RPM por, 15 min, a 4°C. Foram colhidas alíquotas de 500 μL do sobrenadante e transferidas para tubos tipo Ependorf contendo 500 μL do reagente ácido de Salkowski (0,5 M FeCl3, 7,9 M de H2SO4). Após a mistura do reagente e sobrenadante, a mesma foi homogeneizada e colocada ao abrigo da luz por 30 minutos para o desenvolvimento da cor característica do teste. O desenvolvimento da cor rosa indica a produção de IAA qualitativo, a intensidade da coloração rosa é diretamente proporcional à concentração de AIA presente no meio (KUSS et al, 2007). Após esse período procedeu-se a leitura da absorbância em espectrofotômetro a 540 nm. O cálculo da quantidade AIA produzida pelas bactérias foi realizado através do preparo de curva padrão com as concentrações de AIA iguais a 0,1, 0,5, 1,0, 2,0, 5,0, 10,0, 15,0, 20,0 e 30,0 μg.mL-1. Foi obtida a equação C = 13,42.A – 0,338 com coeficiente de correlação igual a 0,997. Onde C é igual a concentração e A igual absorbância.
Resultado e discussão
O método quantitativo de dosagem de AIA se mostrou sensível para o teste
realizado com as cepas isoladas de raízes de arroz e para os controles
testados. Foi observado que todas as cepas analisadas possuem a capacidade
de sintetizar fiotormônios com anel indol em sua cadeia estrutural.
Dentre as cepas analisadas foi observado valores de produção variando entre
0,207 μg.mL-1 a 33,212 μg.mL-1 Figura 1. Dentre as
cepas isoladas as que apresentaram maior produção de AIA foram as ISR82,
ISR5, ISR134, ISR58, ISR73 e ISR30. As estirpes controle de maior produção
foram as BR522 e BR322. Os isolados de menor potencial produtivo do
fitormônio foram ISR80 e ISR65 com concentrações abaixo de 1,0
μg.mL-1.
Os valores obtidos para a concentração de AIA foram divididos em quartis
para classificar de forma semi-quantitativa as bactérias utilizadas no
estudo. Quartis promovem a divisão de um grupo em quatro partes, as classes
definidas como produtores fortes, moderados, com baixa produção ou não
produtores de AIA. Essa ferramenta auxilia na divisão de um grupo maior em
grupos menores, expressando o lugar que um determinado indivíduo ocupa
dentro de seu grupo (BISQUERRA, SARRIERA, MATÍNEZ, 2009).
O primeiro quartil selecionado foi designado para as bactérias não
formadoras de AIA. O segundo quartil foi designado às bactérias de baixa
produção, a este foi atribuído valores iguais à mediana da primeira metade
dos valores ordenados de forma crescente, selecionando valores abaixo de
25%. O ponto de corte para o primeiro quartil foram concentrações iguais ou
menores que 2,851 μg.mL-1.
O quarto quartil foi designado para as cepas com forte produção de AIA. Este
quartil foi determinado pela mediana da segunda metade dos valores ordenados
de forma crescente, selecionando valores acima de 75%. O ponto de corte
para o terceiro quartil foi igual ou maior que 30,387 μg.mL-1. O
terceiro quartil foi ocupado pelas cepas com moderada produção do composto.
Este quartil é a mediana da população, englobando valores 50% abaixo e 50%
acima. Os valores englobados pelo terceiro quartil são valores maiores que
2,851 μg.mL-1 e menores que 30,387 μg.mL-1.
Entre as bactérias testadas são classificadas como produtoras fracas de AIA,
consequentemente se enquadrando no segundo quartil as cepas ISR80, ISR65,
CFN42 e BR10055. Enquadram se no terceiro quartil sendo assim classificadas
como moderadas as cepas ISR59, ISR40, ISR132, ISR31, ISR25, BR520, ISR127,
ISR54, BR522, ISR82 e ISR5. No quarto quartil sendo classificadas como forte
produtoras estão as cepas BR322, ISR134, ISR58, ISR73 e ISR30.
Kuss e colaboradores (2007) avaliando a produção de AIA por bactérias
diazotróficas endofíticas isoladas de cultivares de arroz irrigado,
observaram a produção de AIA entre 2,79 e 13,47 μg.mL-1 após 72
horas de crescimento em meio DYGS sem a adição de triptofano ao meio.
Stroschein e colaboradores (2011) por sua vez avaliando a produção de AIA
por rizóbios isolados de alfafa em meio LM enriquecido com triptofano,
observaram que em todos os isolados testados produziam AIA, com valores que
variaram de 43,04 a 101,26 µg.mL-1.
Shilindwein e colaboradores (2008) em estudos usando quatro isolados de
Bradyhizobium sp e um de Rhizobium leguminosarum observaram
que a produção de AIA apresentau valores significativamente diferentes entre
os gêneros testados. A produção de AIA variou entre 1,2 a 171,1
μg.mL-1 sendo a cepa de Rhizobium legumnosarum o de maior
produção 171,1 μg.mL-1.
No presente trabalho, o valor médio de produção de AIA foi igual a 14,009
μg.mL-1, valor próximo ao obtido por Kuss e colaboradores (2007)
em seus estudos. Em ambos os trabalhos as cepas foram isoladas de plantas de
arroz, podendo indicar desta forma um valor médio de produção de AIA por
estas bactérias quando em simbiose com esta plantas deste gênero.
A seleção de bactérias para produção de AIA em associação com plantas passa
pela avaliação de certos parâmetros, entre eles a capacidade de produção do
composto pela bactéria, a faixa ideal de concentração exigida do hormônio
pela planta e de parâmetros reguladores da produção AIA. O hormônio AIA
assim como os outros fitormônios estimulam o crescimento de plantas apenas
dentro de uma faixa estreita de concentração. Fora da faixa ideal, baixas
concentrações são ineficientes e altas concentrações são tóxicas. Essa faixa
varia entre as diferentes culturas de leguminosas (SCHLINDWEIN et al, 2008).
Em alguns casos, o AIA também pode funcionar na regulação do gene ipdC
através de mecanismo de feedback positivo, fato descrito para
Azospirillum brasilense. (BROEK et al, 1999). Onde o aumento da
concentração de AIA no meio induz a expressão do gene ipdC que codifica que
codifica a enzima indol-3-piruvato descarboxilase. Fato também observado
por SOMERS e colaboradores (2005).
O gene ipdC, também pode ser controlado pela disponibilidade de triptofano
no meio como observado em cepas de Pseudomonas putida GR12-2. Na
rizosfera as bactérias podem obter triptofano a partir do exsudato das
raízes das plantas, assim de certa maneira em alguns casos a planta
hospedeira controla a produção de microbiana de AIA (DUCA et al, 2014).
Outro fator a se observar na seleção de micro-organismos para a produção de
AIA é a via de síntese a qual se utiliza a bactéria. Há evidencias que
micro-organismos podem selecionar uma via metabólica diferenciada,
dependendo do meio ambiente. Fato observado em espécies de Rhizobium
que utilizavam a via do indol-3-acetamida quando associados ao hospedeiro e
a vida do ácido indol-3-piruvato quando em vida livre (RADWAN, MOHAMED,
REIS, 2004).
Estudos como a caracterização fenotípica, ensaios de formação de biofilme,
viabilidade celular, testes de inoculação em leguminosas e outros testes
bioquímicos serão realizados com as cepas descritas neste trabalho, a fim de
selecionar potenciais bactérias promotoras de crescimento vegetal no intuito
de utilizá-las como inoculantes agrícolas.
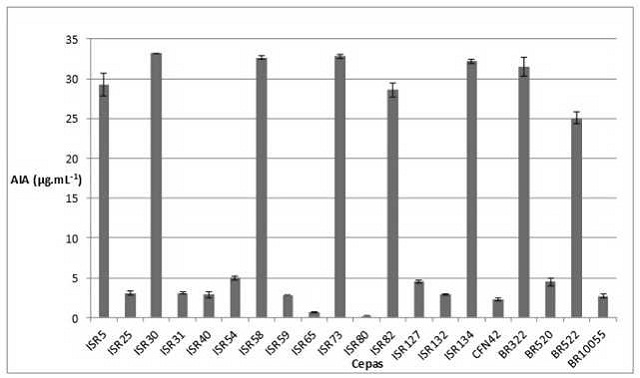
Concentração de AIA produzido pelas cepas em estudo crescidas em meio batata suplementado com triptofano.
Conclusões
Através do experimento realizado observou-se que todas as cepas isoladas dos nódulos de arroz da família Rhizobiaceae, apresentaram mesmo que em baixas quantidades a capacidade de síntese da auxina ácido indol-3-acético a partir do precursor triptofano. Dos isolados 26,7% se mostraram forte produtores, 60% moderados e 13,3% de baixa produção. Entre as cepas controle duas se apresentaram como forte produtoras, uma moderada e duas de baixa produção. O valor médio de produção de AIA pelas cepas neste estudo foi de 14,009 μg.mL-1. Através do presente estudo pode se dizer que todas as cepas possuem potencial como bactérias promotoras de crescimento vegetal, devido a sua capacidade de produção de AIA, podendo ser usadas como alternativa à fertilizantes químicos.
Agradecimentos
A CAPES pela bolsa de mestrado, ao CNPq pelo INCT em Fixação Biológica de Nitrogênio e pelo Programa de Bolsa de Incentivo ao Pesquisador (PROBIP).
Referências
BABALOLA, O. O. Beneficial Bacteria of Agricultural Importance. [b]Biotechnol Lett[/b], n. 32,1559-1570, 2010.
BARTEL, B. Auxin Biosynthesis. [b]Annu. Rev. Plant Physiol. Plant Mol. Biol.[/b] n. 48, 51–66, 1997.
BISQUERRA, B.; SARRIERA, J. C. MATÍNEZ, F. Introdução a Estatística: Enfoque informático com o pacote estatístico SPSS. [b]Bookman[/b], Porto Alegre, 1° Ed, 2009, p. 58.
BONILLA, G. A. E. [i]Seleção de Bactérias Diazotróficas Solubilizadoras de Fósfato e seu Efeito no Desenvolvimento de Plantas de Arroz[/i]. Rio de Janeiro, 86f, 2011. Dissertação (Mestrado em Fitotecnia) Universidade Federal Rural do Rio de Janeiro, 2011.
BRANDLAND, M. T.; LINDOW, S. E. Cloning and Characterization of a Locus Encoding an Indolepyruvate Decarboxylase Involved in Indole-3-Acetic Acid Synthesis in Erwinia herbicola. [b]Applied and Environmental Microbiology[/b], v. 62, n. 11, 4121–4128, 1996.
BROEK, A. V.; LAMBRECHT, M.; EGGERMONT, K.; VANDERLEYDEN, J. Auxins Upregulate Expression of the Indole-3-Pyruvate Decarboxylase Gene in Azospirillum brasilense. [b]J. of Bacteriology[/b], v. 181, n. 4, 1338-1342, 1999.
CHEN, H.; ZHANG Z. X.; ZHANG, G. M.; GUO, X. F.; ZHANG, H. S.; WANG, H. Liquid Chromatographic Determination of Endogenous Phytohormones in Vegetable Samples Based on Chemical Derivatization with 6-Oxy(acetylpiperazine) Fluorescein. [b]J. Agric. Food Chem[/b], v. 58, 4560–4564, 2010.
DIMKPA, C. O.; ZENG, J. MCLEAN, J. E. BRITT, D. W.; ZHAN, J. ANDERSON, A. J. Production of Indole-3-Acetic Acid via the Indole-3-Acetamide Pathway in the Plant-Beneficial Bacterium Pseudomonas chlororaphis O6 Is Inhibited by ZnO Nanoparticles but Enhanced by CuO Nanoparticles. [b]Appl. and Envir, Microb.[/b], v. 78, n. 5, 1404–1410, 2012.
DOBEREINER, J.; ANDRADE, V. O.; BALDANI, V. L. D. Protocolos para Preparo de Meios de Cultura da Embrapa Agrobiologia. Rio de Janeiro, 1999. Disponível em: <http://ainfo.cnptia.embrapa.br/digital/bitstream/CNPAB-2010/27335/1/doc110.pdf> Acesso em: 013 jul. 2015.
DUCA, D.; LORV, J.; PATTEN, C. L.; ROSE, D.; GLICK, B. R. Indole-3-acetic acid in Plant–microbe Interactions. [b]Antonie van Leeuwenhoek[/b], v. 106, 85-125, 2014.
KUSS, A. V.; KUSS, V. V.; LOVATO, T.; FLÔRES, M. L.; Fixação de Nitrogênio e Produção de Ácido Indolacético in vitro por Bactérias Diazotróficas Endofíticas. [b]Pesq. agropec. bras.[/b], v.42, n.10, 1459-1465, 2007.
MANO, Y.; NEMOTO, K. The Pathway of Auxin Biosynthesis in Plants. [b]J. of Experi. Botany[/b], v. 63, n. 8, 2853–2872, 2012.
NETO, D. D.; DARIO, G. J. A.; VIEIRA JÚNIOR, P. A.; MANFRON, P. A.; MARTIN, T. N.; BONNECARRÉRE, R. A. G.; CRESPO, P. E. N. Aplicação e Influência do Fitorregulador no Crescimento das Plantas de Milho. [b]Rev. Da FZVA[/b], v. 11, n. 1, 1-9, 2004.
PRINSEN, E.; CHAUVAUX, N.; SCHMIDT, J.; JHON M.; WIENEKE, U.; GREEF, J.; SCHELL, J.; ONCKELEN, H. V. Stilulation of Indol-3-acetic Production in Rhizobium by Flavonoids. [b]FEBS[/b], v. 282, 53-55, 1991.
RADWAN T. E. S. E. D.; MOHAMED, Z. K.; REIS, V. M.; Aeração e Adição de Sais na Produção de Ácido Indol Acético por Bactérias Diazotróficas. [b]Pesq. agropec. bras.[/b], v.40, n.10, 997-1004, 2005.
SCHLINDWEIN, G.; VARGAS, L. K.; LISBOA, B. B.; AZAMBUJA, A. C.; GRANADA, C. E.; GABIATTI, N. C.; PRATES, F.; STUMPF, R. Influência da Inoculação de Rizóbios Sobre a Germinação e o Vigor de Plântulas de Alface. [b]Ciên. Rural[/b], v. 38, n. 3, 658-664, 2008.
SOMERS, E.; PTACEK, D.; GYSEGOM, P.; SRINIVASAN, M.; VANDERLEYDEN, J. Azospirillum brasilense Produces the Auxin-Like Phenylacetic Acid by Using the Key Enzyme for Indole-3-Acetic Acid Biosynthesis. [b]Applied and Environ. Microbiology[/b], v. 71, n. 4, 1803-1810, 2005.
SPAEPEN, S.; VANDERLEYDEN, J.; REMANS, R. Indole-3-acetic acid in Microbial and Microorganism-Plant Signaling. FEMS Microbiol Rev, v. 31, 425–448, 2007.
STROSCHEIN, M. R. D.; SÁL, E. L. S.; MACHADO, R. G.; CABRAL, T. L.; BRUXEL, M.; GIONGOL, A.; FONTURA, R. C. Caracterização e Influência de Rizóbios Isolados de Alfafa na Germinação e Desenvolvimento Inicial de Plântulas de Arroz. [b]Ciênc. Rural[/b], v. 41, n. 10, 1738-1743, 2011.
SUGAWARAA, S.; HISHIYAMAC, S.; JIKUMARUA, Y.; HANADAA, A. NISHIMURAB, T.; KOSHIBAB, T.; ZHAOD, Y.; KAMIYAA, Y.; KASAHARAA. H. Biochemical Analyses of Indole-3-acetaldoxime Dependent Auxin Biosynthesis in Arabidopsis. [b]PNAS[/b], v. 106, n. 13, 5430-5435, 2009.
SUGAWARA, S.; MASHIGUCHI, K.; TANAKA, K. HISHIYAMA, S.; SAKAI, T.; HANADA, K.; KINOSHITA-TSUJIMURA, K.; YU, H.; DAI, X.; TAKEBAYASHI, Y.; TAKEDA-KAMIYA, N.; KAKIMOTO, T.; KAWAIDE, H.; NATSUME, M.; ESTELLE, M.; ZHAO, Y.; HAYASHI, K. I.; KAMIYA, Y.; KASAHARA, H. Distinct Characteristics of Indole-3-Acetic Acid and Phenylacetic Acid, Two Common Auxins in Plants. [b]Plant Cell Physiol[/b], v. 0, 1-14, 2015.
TSAVKELOVA, E.; OESER, B.; OREN-YOUNG, L.; ISRAELI, M.; SASSON Y.; TUDZYNSKI, B.; SHARON, A. Identification and Functional Characterization of Indole-3-acetamide Mediated IAA Biosynthesis in Plant-Associated Fusarium Species. [b]Fungal Genetics and Biology[/b], n. 49, 48-57, 2012.
Patrocinadores







Apoio









Realização

