ÁREA: Produtos Naturais
TÍTULO: Ação de amostras proteicas/lectina ligadora de quitina da entrecasca de Stryphnodendron adstringens sobre cupins Nasutitermes corniger.
AUTORES: Melo, A.S. (UFPE) ; Silva, D.G.R. (UFPE) ; Pereira, P.R.S. (UFPE) ; Santos, J.M. (UFPE) ; Sá, R.A. (UFPE)
RESUMO: Lectinas apresentam ação contra insetos e fungos. O trabalho objetivou extrair
lectina de Stryphnodendron adstringens, planta do Agreste pernambucano, e
investigar a sua ação sobre os cupins Nasutitermes corniger. 10g de farinha da
entrecasca de S. adstringens foram submetido à extração de proteínas em NaCl 0,15
M, por 8h. A presença de lectina em ES8 foi detectada através da AH. ES8, com AHE
mais elevada, foi escolhida para o processo de purificação parcial. A purificação
foi iniciada com a precipitação de proteínas solúveis no ES8. A fração F4
apresentou a melhor AHE (270034,0), constituindo material adequado para a
purificação da lectina. F4 e ES8 (2,0, 4,0 e 8,0 mg/mL) apresentaram uma eficiente
atividade termiticida para operários e soldados de N. corniger.
PALAVRAS CHAVES: S. adstringens; Térmita; Lectinas
INTRODUÇÃO: A evolução do conhecimento dentro da Bioquímica ecológica tem iluminado os
diversos degraus de complexas interações e adaptações co-evolucionárias que
ocorrem entre plantas, animais e microrganismos. As plantas desenvolveram
mecanismos de respostas e defesa a agressões de predadores. Desta forma, elas
sempre conseguiram dominar as paisagens do planeta (HARBORNE, 1993). A dimensão
dos danos causados por fitopatógenos e pestes de insetos depende do hábito e do
tamanho da população do agente causador e da resistência da planta ao tipo de
ataque (KOGAN, 1994).
Proteínas de plantas que podem estar envolvidas em mecanismos de defesa são as
lectinas, as proteínas inativadoras de ribossomos, os inibidores de proteases e
glicohidrolases(CARLINI & GROSSI-DE-SÁ, 2002; MACEDO et al., 2003).
Stryphnodendron adstringens (barbatimão), tem sua casca amplamente empregada na
medicina popular no tratamento de hemorragias e cicatrização (LORENZI & MATOS,
2002). Extratos hidroalcoólicos de barbatimão apresentaram taninos, alcalóides,
flavonóides e saponinas (SIMÕES et al., 2004).
O termo lectina originado do latim, que significa selecionado, refere-se à
habilidade dessas proteínas ligarem-se seletivamente e reversivelmente a
carboidratos. Em vegetais as lectinas são frequentemente isoladas de sementes e
folhas. Lectinas vegetais podem desempenhar importantes papéis tais como
proteínas de reserva, no mecanismo de defesa contra microrganismos
fitopatogênicos, insetos e animais predadores afetando o crescimento e
desenvolvimento de insetos e apresentando atividades tóxicas em animais
herbívoros (LIS & SHARON, 1986). O trabalho objetivou extrair lectina de
Stryphnodendron adstringens, planta do agreste pernambucano, e investigar a ação
de frações proteicas/lectinas sobre os cupins N. corniger.
MATERIAL E MÉTODOS: 10g de farinha da entrecasca de S. adstringens foram submetido à extração de
proteínas em NaCl 0,15 M, por 8h (ES8).
A atividade lectínica de ES8 foi realizada em placas de microtitulação. Foram
colocados 50µl de NaCl 0,15M em cada poço; em seguida, foram adicionados ao
segundo poço 50 µl da amostra a ser avaliada. Após uma diluição seriada, 50µl de
eritrócitos de coelhos foram adicionados em todos os poços e as placas foram
incubadas por 45 minutos. A AH correspondeu ao inverso da maior diluição (título
-1), na qual ainda se observou a aglutinação total dos eritrócitos.
Esta atividade está relacionada à especificidade e afinidades dos sítios de
ligação das lectinas a carboidratos presentes nas paredes dos eritrócitos.
EB8 foi avaliado quanto à presença de proteínas por espectrofotometria. O método
utilizado foi o de LOWRY et al. (1951). A etapa inicial de purificação foi
realizada com o fracionamento das proteínas de EB8 através da adição de
(NH4)2SO4 segundo GREEN e HUGHES (1955). Em
seguida, o precipitado (F4), foi avaliado na presença de carboidratos.
F4 foi aplicado em coluna de quitina, equilibrada com NaCl 0,15 M. O pico
protéico ativo (PII) foi eluído com Ácido acético 1,0 M. A avaliação da
estabilidade térmica de EB8 e F4 foi efetuada de acordo com SHIOMI et al.
(1979).
A atividade termiticida foi avaliada através de um ensaio “sem escolha” baseado
no método de KANG et al. (1990). A avaliação foi realizada diariamente até a
observação de mortalidade total dos insetos.
RESULTADOS E DISCUSSÃO: A fração F4 apresentou a melhor AHE (270034,0), constituindo material adequado
para a purificação da lectina. AHE foi determinada pela razão AH/concentração de
proteínas (mg/mL).
A AH de F4 foi parcialmente inibida por N-acetilglicosamina. A inibição com N-
acetilglicosamina é um indicativo para purificação da lectina através de
cromatografia em colunas de quitina. F4 foi aplicada em coluna de quitina. O
pico proteico (1,0 mg/mL de proteínas) ativo (PII) eluído com ácido acético 1,0
M foi definido como a lectina de S. adstringens.
A AH de EB8 e F4 (100 μg/mL de cada amostra) foram avaliadas com diferentes
temperaturas e o ensaio indicou que a AH é termoresistente, permanecendo elevada
após aquecimento a 80ºC. Comportamento semelhante foi observado com Cramoll, a
lectina de sementes de Cratylia mollis que, na temperatura de 80°C, manteve AH
elevada (CORREIA & COELHO, 1995).
Lectinas ligadoras de quitina também apresentam ação inseticida para muitos
insetos (CARLINI & GROSSI-DE-SÁ, 2002; MACEDO et al., 2007) e, portanto, o
efeito de ES8 e F4 foram avaliados sobre cupins. ES8 e F4 (2,0, 4,0 e 8,0 mg/mL)
induziram a mortalidade de N. corniger (Figura 1), indicando a presença de
agentes inseticidas.para operários e soldados de N. corniger. A ação tóxica de
lectinas inseticidas tem sido relacionada à ligação dessas proteínas a
glicoproteínas presentes no trato digestivo de insetos. Lectinas entomotóxicas
são resistentes à degradação proteolítica no intestino de insetos e podem se
ligar à membrana peritrófica na região do intestino médio, interferindo na ação
de enzimas digestivas e proteínas de assimilação (MACEDO et al., 2002).
Figura 1 - Sobrevivência de insetos de N. corniger em contato com o Ex
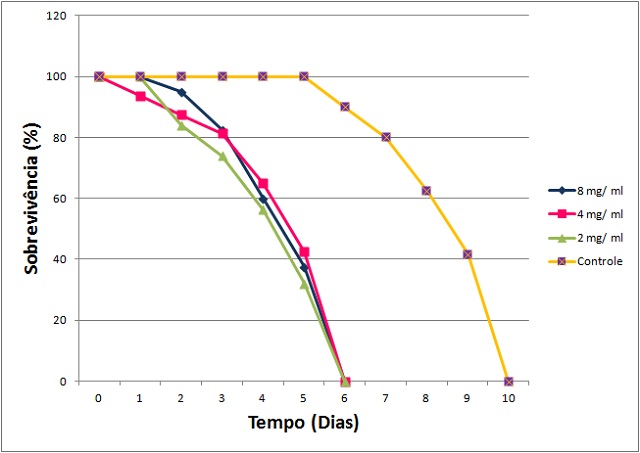
Efeito do EB8 em contato com insetos de N. corniger.
Dosagens utilizadas: 2,0; 4,0 e 8,0 mg/mL; além de
NaCl 0,15 M como controle negativo.
Figura 2 - Sobrevivência dos insetos de N. corniger em contato com a F
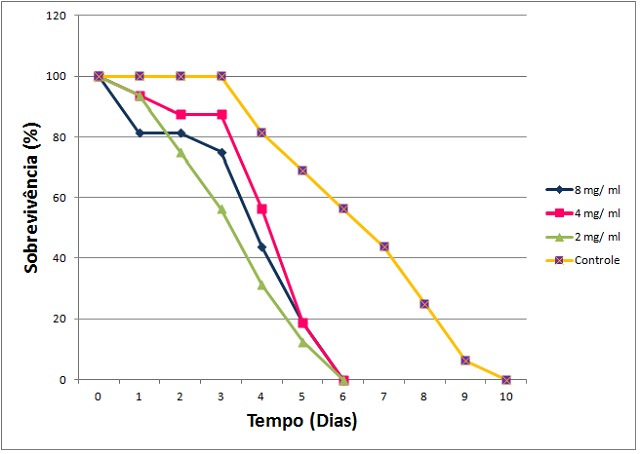
Efeito da F4 em contato com insetos de N. corniger.
Dosagens utilizadas: 2,0; 4,0 e 8,0 mg/mL; NaCl 0,15
M como controle negativo.
CONCLUSÕES: A fração proteica foi inibida com N-acetilglicosamina, que estimulou a purificação
da lectina (PII) em cromatografia de matriz quitina. As frações proteicas EB8 e F4
mostraram-se termoresistentes, mantendo sua AH acima de 80º C. ES8 e F4
apresentaram ação inseticida sobre cupins de Nasutitermes corniger.
AGRADECIMENTOS: À Profª Drª Patrícia M. G. Paiva, da UFPE-Campus Recife; e ao Dr. Thiago
H. Napoleão, integrante da mesma unidade; pelo apoio incondicional a mim dado.
REFERÊNCIAS BIBLIOGRÁFICA: CARLINI, C. R., GROSSI-DE-SÁ, M. F. Plant toxic proteins with insecticidal properties. A review on their potential as bioinsecticides. Toxicon 40: 1515–1539, 2002.
CORREIA, M. T. S.; COELHO, L. C. B. B. Purification of a glucose/mannose specific lectin, isoform 1, from seeds of Cratylia mollis Mart. (camaratu bean). Applied Biochemistry and Biotechnology 55: 261-273, 1995.
LOWRY, O.H. et al. Protein measurement with the Folin phenol reagent. Journal of Biological Chemistry 193, 265–275, 1951.
GREEN, A. A.; HUGHES, W. L. Protein fractionation on the basis of solubility in aqueous solutions of salts and organic solvents. In: COLOWICK, S.; KAPLAN, N. Methods in Enzymology, New York: Academic Press, v.1, pp. 67-90, 1995.
HARBORNE, J. B. Introduction to ecological biochemistry. 4ª ed. London: Academic Press, 1993.
KANG, H. Y.; MATSUSHIMA, N.; SAMESHIMA, K.; TAKAMURA, N. Termite resistance tests of hardwoods of Kochi growth. The strong termiticidal activity of kagonoki (Litsea coreana L´eveill´e). Mokuzai Gakkaishi 36: 78–84, 1990.
KOGAN, M.Plant resistance in pest management. In: METCALF, R. L; luckmann, w, h. Introduction to insect pest management. New York: Wiley –Interscience Publication, 1994.pp. 103-146.
LIS, H.; SHARON, N. Biological properties of lectins. In: LIENER, I. E.; SHARON, N.; GOLDSTEIN, I. J. (eds). The Lectins: Properties, Functions, and Applications in Biology and Medicine, New York: Academic Press. pp. 265-291, 1986.
LORENZI, H.; MATOS, F. J. A. Plantas Medicinais do Brasil Nativas e Exóticas.
MACEDO, M. L. R.; FREIRE, M. G. M.; NOVELLO, J. C.; MARANGONI S. Talisia esculenta lectin and larval development of Callosobruchus maculatus and Zabrotes subfasciatus (Coleoptera: Bruchidae). Biochimica et Biophysica Acta 1571: 83–88, 2002.
MACEDO, M. L. R.; FREIRE, M. G. M.; SILVA, M. B. R.; COELHO, L. C. B. B. Insecticidal action of Bauhinia monandra leaf lectin (BmoLL) against Anagasta kuehniella (Lepidoptera: Pyralidae), Zabrotes subfasciatus and Callosobruchus maculatus (Coleoptera: Bruchidae). Comparative Biochemistry and Physiology A 146: 486-498, 2007.
SHIOMI, K.; KAMIYA, H.; SHIMIZU, Y. Purification and characterization of an agglutinin in the red alga Agardhiella tenera. Biochimica et Biophysica Acta 576: 118-127, 1979.
SIMÕES, CM.O.; SCHEMKEL, E. P.; GOSMANM, G.; MELO, J. C. P.; MENTZ, L.; SOUSA, C.R., “Contribuição ao Conhecimento Químico de Plantas do Ceará, Fortaleza. 1999.
