Otimização da produção de proteases por Penicillium citrinum
ISBN 978-85-85905-23-1
Área
Bioquímica e Biotecnologia
Autores
Santos, G.L. (UNIVERSIDADE ESTADUAL DE CAMPINAS) ; Melani, N.B. (UNIVERSIDADE ESTADUAL DE CAMPINAS) ; Campos, E.S. (UNIVERSIDADE FEDERAL DE UBERLÂNDIA) ; Tambourgi, E.B. (UNIVERSIDADE ESTADUAL DE CAMPINAS)
Resumo
Embora as proteases sejam enzimas de grande importância no mercado atual, a maioria destas são importadas. Desse modo, se faz necessário o aumento da produção nacional. O objetivo deste trabalho foi de avaliar meios para produção de proteases por Penicillium citrinum através de fermentação submersa. A fim de se obter o melhor meio fermentativo, foi analisado o efeito de diferentes tipos de fontes de carbono e de nitrogênio na produção. Assim, foram determinadas as atividades proteolíticas através do uso de azocaseína como substrato. O valor mais baixo foi de 0,18 U/mL, exibido pelo meio contendo glicose e peptona. Foi verificado que o meio contendo glicose e extrato de levedura exibiu a maior produção e se mostrou indutor de protease, com uma atividade proteolítica de 2,08 U/mL.
Palavras chaves
proteases; fermentação; Penicillium citrinum
Introdução
No cenário atual, as enzimas apresentam importante papel na indústria, sendo empregadas em processos de diversos setores como, por exemplo, de alimentos, têxtil, química, de papel, entre outros. Dentre essas enzimas, se encontram as proteases. As proteases, também denominadas enzimas proteolíticas, apresentam a capacidade de hidrolisar pequenos peptídeos. Quanto à classificação destas enzimas, segundo o Comitê de Nomenclatura Enzimática (EC) da União Internacional de Bioquímica e Biologia Molecular, estas pertencem à classe 3 (hidrolases) e subclasse 3.4 (peptidases). Essas biomoléculas podem ser classificadas quanto a sua região de atuação nas cadeias polipeptídicas: as peptidases que clivam ligações no interior da cadeia são classificadas como endopeptidases (EC 3.4.21-99), enquanto as que atuam nas extremidades são chamadas de exopeptidases (EC 3.4.11-19) (BON et al., 2008). Nos seres humanos, as proteases estão envolvidas com processos biológicos essenciais, como a coagulação sanguínea, a diferenciação dos tecidos e a morte celular. Além disso, essas biomoléculas apresentam diversas aplicações, encontrando-se entre as mais importantes classes de enzimas industriais (WISEMAN, 1991). Estas podem ser usadas na indústria farmacêutica, de couro, no processamento de alimentos, na fabricação de detergentes, na produção de bebidas, no aumento da digestibilidade em alimentos de animais, entre outros. As enzimas proteolíticas podem ser produzidas por plantas, animais, e por microrganismos. Dentro desta última categoria se destacam os vírus, fungos e bactérias. Devido a algumas características, as proteases microbianas se mostram bem vantajosas, pois apresentam fonte de rápido crescimento, possibilidade de cultivo em espaço limitado e fácil manipulação genética (TREMACOLDI, 2009). Além disso, apresentam maior estabilidade que suas homólogas animais e de plantas (WISEMAN, 1991). Cerca de 40% das proteases utilizadas industrialmente são de origem microbiana (JOHNVESLY, NAIK; 2001). Dentre os microrganismos, os fungos são os que apresentam maior variedade de proteases, sendo que a produção comercial se dá principalmente pelos gêneros Aspergillus e Penicillium (COLEN, 2006). Essas biomoléculas apresentam grande destaque, correspondendo a mais de 65% do mercado total de enzimas (ANNAMALAI et al., 2014). No Brasil, a maioria dessas enzimas é importada, já que sua produção no país é muito baixa e representa menos de 2% do total (GIONGO, 2006), se fazendo necessário o crescimento da produção nacional. Dadas as circunstâncias e tendo em vista das vantagens apresentadas pelas enzimas microbianas, o presente trabalho tem como objetivo a produção otimizada de proteases através de Penicillium citrinum.
Material e métodos
Microrganismo e Conservação O fungo filamentoso Penicillium citrinum foi cedido pelo Departamento de Microbiologia e Parasitologia, da Universidade Federal de Pernambuco. O microrganismo foi inserido em erlenmeyers contendo caldo Sabouraud (peptona 10 g/L e glicose 40 g/L), o qual foi cultivado por 120 horas, sob uma agitação em agitador orbital de 150 rpm à 28 ºC, sendo armazenado posteriormente. Produção Proteolítica e Extrato Enzimático O pré-inóculo foi fermentado em meio Sabouraud, o qual contêm peptona 10 g/L e glicose 40 g/L por 120 horas, 150 rpm, a 28º C. Visando a produção de protease, foi avaliado o melhor meio de fermentação contendo diversas fontes de carbono e nitrogênio. Esse processo ocorreu através de uma fermentação submersa de 240 horas, agitada a 150 rpm e a 28º C. As fontes de carbono se encontram na concentração de 10 g/L, enquanto que as de nitrogênio de 2g/L. Em cada um dos meios foi inoculado cerca de 106 esporos. Cada teste foi realizado em triplicata e, assim, ao final de cada processo fermentativo, foi avaliada a atividade proteolítica e a quantidade de proteínas totais. Determinação da Atividade Proteolítica A atividade proteolítica foi determinada de acordo com a metodologia descrita por Charney e Tomarelli (1947) através do uso do substrato azocaseína (Sigma, St. Louis, MO, USA). Dessa forma, 125 µL do extrato enzimático reagiram com 125 µL de azocaseína a 1% em solução tampão fosfato de sódio 0,1 M, pH 7,0, durante 15 min a 37 ºC. Em seguida, a reação foi finalizada com a adição de 750 µL de ácido tricloroacético (TCA) 5% e, uma centrifugação a 6.000 g por 5 min a 25 °C. Assim, foi acrescentado 1 mL de hidróxido de sódio (NaOH) 0,5 M, ao sobrenadante. As leituras da absorbância à 440 nm foram feitas, utilizando água destilada como amostra branco. A atividade proteolítica é expressa em unidades de atividade enzimática (U), em que uma unidade equivale a uma quantidade de enzimas capazes de aumentar em 1 a absorbância por hora, nestas condições. Determinação de Proteínas Totais Para a determinação da concentração de proteínas totais, foi utilizado o método descrito por Bradford (1976), em que ocorre uma reação colorimétrica entre grupos aromáticos da proteína e o corante Coomassie brilliant blue em meio ácido. Após a centrifugação do meio fermentativo, 20 µL do extrato enzimático e 1 mL do reagente de Bradford foram inseridos em microtubos, sendo realizada uma agitação por inversão. Em seguida, estes foram incubados por 5 minutos à temperatura ambiente. Assim, a leitura em espectrofotômetro foi realizada a 595 nm. Uma curva padrão foi previamente construída utilizando como proteína padrão a albumina de soro bovino (BSA).
Resultado e discussão
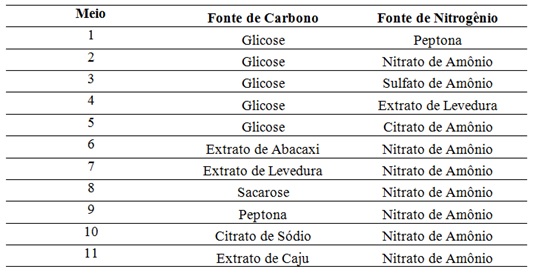
A indução ou inibição da produção de proteases ocorre de acordo com a
composição do meio, sendo que as fontes de carbono e de nitrogênio presentes
neste exercem efeitos reguladores sobre a síntese desta enzima. Desse modo,
as fontes de carbono e nitrogênio avaliadas se encontram apresentadas na
Tabela 1.
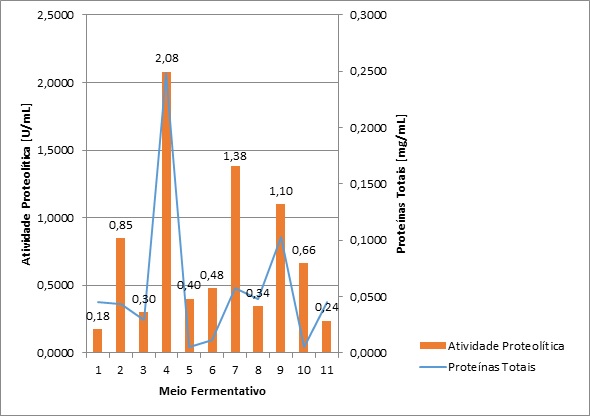
Assim, ao fim dos experimentos, foram efetuados os testes. Realizando a
média entre as triplicatas, os resultados em cada uma das amostras foram
obtidos. Estes se encontram representados no Gráfico 1. Neste, as proteínas
totais são expressas em mg/mL, enquanto que a atividade proteolítica em
U/mL.
Analisando as quantidades de proteínas totais e atividades proteolíticas
obtidas no Gráfico 1, é possível perceber que a síntese destas enzimas esta
relacionada com a produção de biomassa do fungo.
É possível verificar que o meio 1, composto por glicose e peptona, apresenta
o menor valor de atividade proteolítica, equivalente a 0,18 U/mL. Esta
informação concorda com Laxman et al. (2005), que afirma que a
peptona não é boa fonte de nitrogênio para a indução de protease. Outros
valores baixos de produção são encontrados nos meios 3 e 5, que também
apresentam a glicose como fonte de carbono. Levando em consideração que a
glicose é uma das melhores fontes de carbono para a produção de protease
(LAXMAN et al., 2005), esses dados mostram que o sulfato de amônio e
o citrato de amônio não apresentaram efeito positivo na produção desta
enzima. Outros valores baixos de atividade são encontrados nos meios 6, 8,
10 e 11.
O maior valor de atividade é encontrado no meio 4, composto de glicose e
extrato de levedura. Este apresentou uma atividade proteolítica de 2,08
U/mL. Este resultado obtido concorda com Haddar et al. (2010 ) e
Laxman et al. (2005), que demonstraram que o extrato de levedura é
uma das melhores fontes de nitrogênio para a indução da produção de
protease. O meio 7 também se mostrou favorável, apresentando uma atividade
de 0,48 U/ml. Os meios 9 e 2 apresentaram valores semelhantes, com 1,10 e
0,85 U/mL, respectivamente.
Estudos realizados por Chu (1992) e Laxman et al. (2005) mostram que,
quando inseridos nos meios, os sais de amônio se apresentam como inibidores
de produção de protease. Essa informação não concorda com os resultados
apresentados pelos meios 7 e 9, que contêm nitrato de amônio em sua
composição e apresentaram altos valores obtidos.
Conclusões
Tendo em vista os resultados obtidos, o meio contendo glicose e extrato de levedura foi o que apresentou maior produção de protease por fermentação submersa de Penicillium citrinum, sendo então classificados como as melhores fontes de carbono e nitrogênio. Além disso, o nitrato de amônio também se mostrou positivo na indução de produção da enzima. O citrato de amônio, sulfato de amônio e a peptona se mostraram inibidores.
Agradecimentos
O presente trabalho foi realizado com apoio do CNPq, Conselho Nacional de Desenvolvimento Científico e Tecnológico – Brasil.
Referências
ANNAMALAI, N.; RAJESWARI, M. V.; BALASUBRAMANIAN, T. Extraction, purification and application of thermostable and halostable alkaline protease from Bacillus alveayuensis CAS 5 using marine wastes. Food and Bioproducts Processing, v. 92, n. 4, p. 335-342, 2014.
BON, E. P. S.; FERRARA, M. A.; CORVO, M. L. Enzimas em Biotecnologia: Produção, Aplicações e Mercado. Rio de Janeiro: Editora Interciência, 2008, 506 p.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, v. 72, n. 1, p. 248-254, 1976.
CHARNEY, J.; TOMARELLI, R. M. A colonmetric method for the determination of the proteolytic activity of duodenal juice. Journal of Biological Chemistry, v. 171, p. 501-505, 1947.
COLEN, G. Isolamento e seleção de fungos filamentosos produtores de lipases. Faculdade de Farmácia da UFMG, (Belo Horizonte-MG), 2006 [Tese de Doutorado].
GIONGO, J. L. Caracterização e aplicação de proteases produzidas por linhagens de Bacillus sp. 2006. 95 f. Dissertação (Mestrado em Microbiologia Agrícola e do Ambiente) – Faculdade de Agronomia, Universidade Federal do Rio Grande do Sul, Porto Alegre. 2006.
HADDAR, A. et al. Low-cost fermentation medium for alkaline protease production by Bacillus mojavensis A21 using hulled grain of wheat and sardinella peptone. J Biosci Bioeng, v. 110, n. 3, p. 288-294, 2010.
JOHNVESLY, B.; NAIK, G. R. Studies on production of thermostable alkaline protease from thermophilic and alkaliphilic Bacillus sp. JB-99 in a chemically defined medium. Process Biochemistry, Oxford, UK, v. 37, p. 139-144, 2001.
LAXMAN, R. S. et al. Optimization and scale up of production of alkaline protease from Conidiobolus coronatus. Process Biochem, v. 40, p. 3152-3158, 2005.
TREMACOLDI, C. R. Proteases e inibidores de proteases na defesa de plantas contra pragas. 1 ed. Belém, 2009, 44 p.
WISEMAN, A. Manual de biotecnologia de las enzimas. Zaragoza: Acribia, 1991, 444 p.










