ESTUDO IN SILICO DA ATIVIDADE ANTIPARASITARIA DO SCHISTOSSOMA MANSSONI POR MEIO DO COMPMEXO DI-L-CLORO-BIS [CLORO (4,7-DIMETIL-1, 10-FENANTROLINA) CÁDMIO (II)]
ISBN 978-85-85905-25-5
Área
Bioquímica e Biotecnologia
Autores
Sousa, M.B. (UFMA) ; Bastos, R.S. (UFMA-CIENATEC) ; Ribeiro, F.V.S. (IFPA) ; Barbosa, E.S. (UFPA) ; Faria, M.B. (UFMA) ; Silva, W.F. (UFMA) ; Silva, S.T.P. (IFPA) ; Lima, B.A.V. (UFMA-CIENATEC) ; Rocha, J.A. (UFMA-CIENATEC)
Resumo
O Schistosoma mansoni é o causador da Esquistossomose humana uma doença tropical que infecta mais de 300 milhões de pessoas no mundo, apesar isso essa doença ainda é considerada negligenciada. O objetivo deste trabalho é estudar as interações do cádmio com sete alvos enzimáticos da esquistossomose, visando o estudo da sua atividade inibitória in silico através de docagem molecular. Dentre os valores analisados obtivemos uma maior energia de ligação com a PNP, tendo um valor de -7,35 Kcal.mol-1. A maior constante de inibição foi a UP, que obteve 28.05 uM. A partir dos resultados obtidos pôde-se observar a eficiência do ligante na interação com algumas das enzimas parasitarias, com isso o composto torna-se uma opção para o estudo de desenvolvimento de um possível fármaco.
Palavras chaves
Docagem Molecular; Cádmio; Schistossoma manssoni
Introdução
O Schistosoma mansoni causador da Esquistossomose humana é um parasito platelminto trematódeo, sua infecção nos seres humanos acontece em sua maioria por falta de saneamento adequado e a higiene precária nas populações mais pobres (PHILIPPSEN & DEMARCO, 2019), ocorrendo na maioria dos países menos desenvolvidos, é uma doença tropical que apresenta casos de aproximadamente 300 milhões de pessoas infectadas no mundo, apesar da grande quantidade de pessoas ainda é uma doença considerada negligenciada (ROMANELLO et al., 2019). No Brasil ocorre na região Nordeste e no estado de Minas Gerais, sendo relatado cerca de seis milhões de pessoas infectadas pelo parasito (KATZ N. e ALMEIDA K., 2003). No Brasil, foram identificados três tipos de espécie de caramujos responsáveis por abrigar o hospedeiro, são eles: B. glabrata; B. tenagophila; e B. stramínea. Esses moluscos são abundantes em locais de água doce, como por exemplo, barragens, áreas de irrigação, rios, lagos e lagoas (VITORINO RR, et al., 2012). O objetivo deste trabalho é estudar as interações do ligante complexo dimérico di-l-cloro-bis [cloro (4,7-dimetil-1, 10-fenantrolina) cádmio (II)] (WARAD et al. 2012) e sete alvos enzimáticos da esquistossomose, Arginase (ARG), Cathepsin B1 Isotype 1 (2CB1), Histone Deacetylase 8 (HDAC8), Methylthioadenosine Phosphorylase (MTAP), Puntative Uridine Phosphorylase (UP), Purine Nucleoside Phosphorylase (PNP), Thioredoxin Glutathione Reductase (TGR), de códigos respectivos 4Q3P, 3QSD, 4CQF, 4L5A, 4TXH, 1TCU, 2V6O, disponíveis no banco de dados Protein Data Bank (PDB) visando o estudo da sua atividade inibitória por meio de docagem molecular.
Material e métodos
Para fazer a docagem, primeiro foi obtido os desenhos estruturais do ligante com o auxílio do software gráfico GaussView5 e as enzimas referidas. Tendo o desenho da estrutura molecular do ligante cádmio foi realizado a otimização do mesmo, os cálculos computacionais para a otimização foram feitos através do programa Gaussian 9w (Gaussian, 2009), O método adotado foi o Density Functional Theory (DFT), com funcional hibrido B3LYP e conjunto de base genérico, sendo a base 6- 311++G** para os átomos de C, H, N e Cl e a base LANL2DZ para o átomo de cádmio (KLAI et al., 2017; SOUDANI et al., 2016). Os cálculos biológicos do ligante- enzima ocorreram através do software AutoDockTools, preparando o ligante e as macromoléculas para interação, foi ajustado o grid box 60x60x60 no sitio ativo de cada enzima estudada. Para a referida interação foi feita uma alteração nos parâmetros de docagem, adicionando o cloro é cádmio para fazerem os respectivos testes com o ligante (di-l-cloro-bis [cloro (4,7-dimetil-1, 10-fenantrolina)) com as enzimas de schistossoma mansoni: ARG, MTAP, TGR, PNP, HDAC8, 2CB1 e UP. O algoritmo atribuído foi o Lamarckian GA, os demais parâmetros adicionados para a interação foram realizados como o padrão, nas simulações ocorreram 100 corridas independentes, para realizar a análise da menor energia de ligação e a taxa de inibição do ligante complexo dimérico di-l-cloro-bis [cloro (4,7-dimetil- 1, 10-fenantrolina) para cada uma das enzimas do schistossoma (ROCHA et al., 2018; FERREIRA, 2015). O que em seguida sugeria a maior taxa de inibição em cada uma delas de acordo com os cálculos computacionais citados a cima.
Resultado e discussão
A maior energia de ligação ocorreu com a PNP, tendo um valor de -7,35 Kcal.mol-1
(Tabela 1), a MTAP que obteve um resultado de -7,18 Kcal.mol-1, a TGR que obteve
o terceiro maior valor de ligação, elas obtiveram o mesmo número de com formações
no cluster. A enzima que obteve o maior número de ligações hidrofóbicas foi a
ARG, com 16 interações. Nas conformações de interação hidrofóbica a PNP formou 13
e uma ponte de hidrogênio no aminoácido TYR90, se tornando assim não muito forte.
Com esse número de ligações hidrofóbicas empata com a TGR que também formou treze
interações hidrofóbicas e nenhuma ponte de hidrogênio, como vemos na tabela
abaixo. O alvo com a maior constante de inibição foi a UP, que obteve 28,05 uM.
O resultado da interação e ação inibitória do ligante com o alvo 2CB1 obteve o
valor de energético de ligação -6.65 Kcal.mol-¹ formando 11 interações
hidrofóbicas, e nenhuma Ponte de hidrogênio, obtendo uma constante de inibição de
15,78 uM. A energia de ligação com o alvo ARG foi de 264,19 Kcal.mol-1
demonstrando a necessidade de bastante energia para se ligar, ela não obteve
nenhuma taxa de inibição, formou 5 pontes de hidrogênio16 interações
hidrofóbicas.
Ação inibitória cádmio com o alvo UP, formou apenas uma ponte de hidrogênio, e
seis interações hidrofóbicas, obteve o valor energético de ligação de -6,65
kcal.mol-1 e 28,05 uM em sua constante de inibição. A interação do HDAC8 teve uma
energia de ligação de -6,8 Kcal.mol-1 e uma constante de inibição de 10,3 uM
tendo apenas interações hidrofóbicas. O alvo TGR teve uma energia de ligação de
-6,94 Kcal.mol-1 e uma taxa inibitória de 8,24 uM, não houve formação de pontes
de hidrogênio.
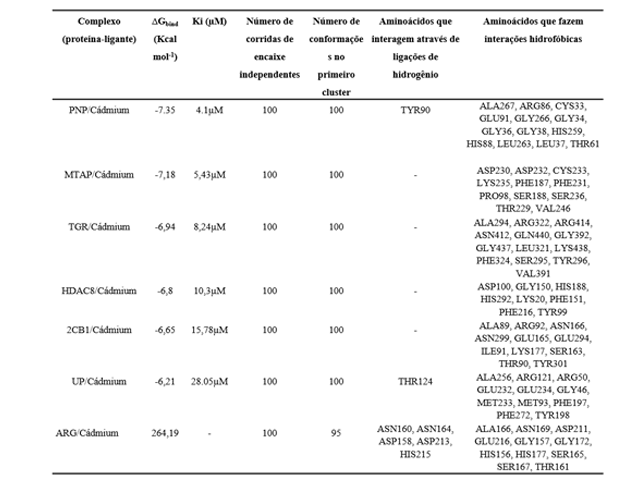
Parâmetros de Docagem Molecular
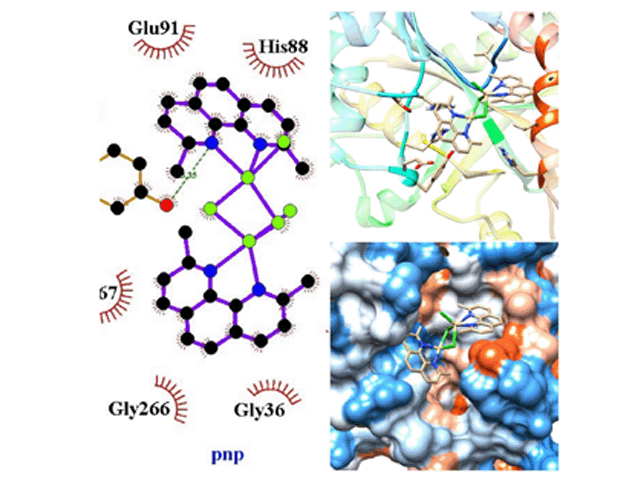
Interação PNP/cádmio, interação em duas dimensões, três dimensões e superfície de contato.
Conclusões
A partir dos resultados obtidos pôde-se observar potencial do composto dimérico di- l-cloro-bis [cloro (4,7-dimetil-1, 10-fenantrolina) cádmio (II)] na interação com algumas das enzimas do parasito de schistossoma mansoni tendo resultado de interações satisfatórios, foi obtido valores favoráveis para a continuação da pesquisa em forma experimental in vitro, valor de maior interação que foi de -7,35 Kcal.mol-1, obtido pela enzima de código PNP. Tornado assim um possível candidato a fármaco antiparasitários para o tratamento das doenças causadas pelo Schistosoma mansoni.
Agradecimentos
Primeiramente agradeço a Deus que sempre esteve me protegendo, agradeço a minha mãe que sempre esteve ao meu lado e a minha família que também me ajuda quando pode. Agradeço ao grupo CIENATEC.
Referências
Frisch, M. J.; Trucks, G. W.; Schlegel, H. B.; Scuseria, G. E.; Robb, M. A.; Cheeseman, J. R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G. A.; Nakatsuji, H.; Caricato, M.; Li, X.; Hratchian, H. P.; Izmaylov, A. F.; Bloino, J.; Zheng, G.; Sonnenberg, J. L.; Hada, M.; Ehara, M.; Toyota, K.; Fukuda, R.; Hasegawa, J.; Ishida, M.; Nakajima, T.; Honda, Y.; Kitao, O.; Nakai, H.; Vreven, T.; Montgomery, J. A., Jr.; Peralta, J. E.; Ogliaro, F.; Bearpark, M.; Heyd, J. J.; Brothers, E.; Kudin, K. N.; Staroverov, V. N.; Kobayashi, R.; Normand, J.; Raghavachari, K.; Rendell, A.; Burant, J. C.; Iyengar, S. S.; Tomasi, J.; Cossi, M.; Rega, N.; Millam, M. J.; Klene, M.; Knox, J. E.; Cross, J. B.; Bakken, V.; Adamo, C.; Jaramillo, J.; Gomperts, R.; Stratmann, R. E.; Yazyev, O.; Austin, A. J.; Cammi, R.; Pomelli, C.; Ochterski, J. W.; Martin, R. L.; Morokuma, K.; Zakrzewski, V. G.; Voth, G. A.; Salvador, P.; Dannenberg, J. J.; Dapprich, S.; Daniels, A. D.; Farkas, Ö.; Foresman, J. B.; Ortiz, J. V.; Cioslowski, J.; Fox, D. J. Gaussian 09, Revision D.01. Gaussian, Inc.: Wallingford CT, 2009.
KATZ N, ALMEIDA K. Esquistossomose, xistosa, barriga d'água. Ciência e Cultura, 2003; 55(1): 38-43.
KLAI, Kacem et al. A Hirshfeld surface analysis, crystalstructureandphysicochemicalstudies of a new Cd (II) complexwiththe 2-amino-4-methylpyrimidine ligand. Journal of Molecular Structure, v. 1128, p. 378-384, 2017.
PHILIPPSEN, Gisele S.; DEMARCO, Ricardo. Impact of transposable elements in the architecture of genes of the human parasite Schistosoma mansoni. Molecular and biochemical parasitology, v. 228, p. 27-31, 2019.
ROCHA, Jefferson A. et al. Computational quantum chemistry, molecular docking, and ADMET predictions of imidazole alkaloids of Pilocarpus microphyllus with schistosomicidal properties. PloS one, v. 13, n. 6, p. e0198476, 2018.
ROMANELLO, Larissa et al. In vitro and in vivo characterization of the multiple isoforms of Schistosoma mansoni hypoxanthine-guanine phosphoribosyltransferases. Molecular and biochemical parasitology, v. 229, p. 24-34, 2019.
SOUDANI, S. et al. A hirshfeld surface analysis, crystalstructureandphysicochemicalcharacterization of 1-ethylpiperazinium trichlorocadmate (II). SolidStateSciences, v. 57, p. 49-55, 2016.
VITORINO RR, et al. Esquistossomose mansônica: diagnóstico, tratamento, epidemiologia, profilaxia e controle. Ver. Soc. Bras. Clín. Med, 2012; 10(1): 39-45.
WARAD, Ismail et al. Novel di-μ-chloro-bis [chloro (4, 7-dimethyl-1, 10-phenanthroline) cadmium (II)] dimer complex: synthesis, spectral, thermal, and crystal structure studies. Research on Chemical Intermediates, v. 39, n. 6, p. 2451-2461, 2013.








