ESTUDO DE DOCKING MOLECULAR DA PIPERINA E METABÓLITOS COM OS RECEPTORES HOMONIO JUVENIL E ACETILCOLINESTERASE RELACIONADOS AO AEDES AEGYPTI
ISBN 978-85-85905-25-5
Área
Bioquímica e Biotecnologia
Autores
Oliveira de Aguiar, C.P. (UFPA) ; Luis Belém dos Santos, K. (UNIFAP/UFPA) ; Carvalho Lobato, C. (UNIFAP/UFPA) ; Marques de Oliveira, H. (ABQ-AP/SEED-AP) ; dos Santos Borges, R. (UFPA)
Resumo
A piperina é a principal amida alcaloide extraída da espécie Piper nigrum Linn, responsável por diferentes propriedades farmacológicas como antinflamatória, antimicrobiana, hepatoprotetora e antioxidante, e tem sido fonte de inspiração para modelagem molecular com finalidade de obter fármacos ou produtos com melhor atividade e menor toxicidade. O bioativo tem se mostrando uma ótima alternativa para o combate ao Aedes aegypti. Estudos de ancoragem molecular foram empregados com o objetivo de identificar as estruturas (piperina e metabólitos) responsáveis pela atividade inseticida. Os resultados da afinidade de ligação demonstraram que os metabólitos M1, M2, M7 e M9 possuem valores superiores ao piperina e são os prováveis responsáveis pela atividade adulticida para o Aedes aegypti.
Palavras chaves
Docking molecular; Estudo in silico; Predição de Compostos
Introdução
A dengue é uma doença causada por arbovírus (DENV), da família Flaviviridae, gênero Flavivírus, e apresenta quatro sorotipos (DENV-1, DENV-2, DENV-3 e DENV-4) que variam clinicamente desde formas autolimitadas a severas, ou até mesmo fatais (RAMOS-CASTAÑEDA, 2017). O mosquito Aedes (Stegomyia) aegypti (LINNAEUS, 1762), pertencente à família Culicidae, ordem díptera, é o principal vetor da doença, além de outras como zika, chikungunya e febre amarela (GIACOPPO et al., 2017). O Brasil está entre os países com maior distribuição global da doença, liderando a américa desde 1995 com 85,7% (1.345.994) dos casos notificados até o primeiro semestre de 2019 (PAHO, 2019). O controle e combate as formas larvais e adultas do Ae. aegypti incluem a gestão ambiental e o controle mecânico, biológico ou químico, dentre outras (ROIZ et al., 2018; ZARA et al., 2016). O emprego de inseticidas químicos ainda é o método mais efetivo no combate as formas do vetor, mas que devido ao seu manejo excessivo tem favorecido o surgimento de resistência das espécies alvo ao longo do tempo (HAMID et al., 2018). A piperina é a principal amida extraída da espécie Piper nigrum Linn, responsável por diferentes propriedades farmacológicas como antinflamatória, antimicrobiana, hepatoprotetora, antimetastásica, antioxidante, além de potenciar a ação de diferentes drogas (GORGANI et al., 2017). O bioativo tem se mostrando uma ótima alternativa para o combate ao mosquito Ae. aegypti por possuir atividade larvicida (MARQUES; KAPLAN, 2015). Neste contexto, os compostos produzidos pelo metabolismo da piperina podem estar envolvidos na atividade relatada. Assim, o presente trabalho visa o estudo teórico da capacidade inseticida de metabólitos da piperina contra as formas imaturas e adultas do Aedes aegypti.
Material e métodos
O total de 9 metabólitos foram obtidos por predição do metabolismo da piperina pelo programa PreADMET (VENKATESH et al., 2017). Posteriormente, todas as estruturas foram desenhadas no programa ChemDraw e otimizadas no programa HyperChem pelos métodos de mecânica molecular (MM+) e semi-empírico (PM3) (HYPERCUB INC., 2009; CHEMOFFICE, 2005). As estruturas da piperina e dos metabólitos são mostradas na Figura 1. A estrutura cristalográfica da piperina foi encontrada no Cambridge Crystallographic Data Center - CCDC (GROOM et al., 2016). Por sua vez, a estrutura cristalográfica da enzima acetilcolinesterase (AChE) de Drosophila melanogaster (KROUPOVA et al., 2018) e para o hormônio juvenil foram obtidas no Banco de Dados de Proteína (PDB), com as respectivas ID 1QON e ID 5V13, respectivamente, pela semelhança na sequência de aminoácidos com o mosquito Ae. aegypti (ENGDAHL et al., 2015; BOTAS, 2017; BLENAU; RADEMACHER; BAUMANN, 2012). As estruturas da proteína e ligantes foram preparadas através do software livre Discovery Studio (DS) Visualizer v.17.2.016349 e salvas no formato PDB (BIOVIA, 2017). O estudo de simulação de acoplamento molecular dos ligantes com a enzima acetilcolinesterase (AChE) foram verificados utilizando o AutoDock Vina 1.1.2 (TROTT, OLSON, 2010), através da interface com o software PyRx 0.8.30, sendo que a validação da ancoragem foi efetuada pela comparação das conformações dos metabólitos inibidores acoplados ao receptor com a proteína inibidora AChE cristalografada (COSTA et al., 2019).
Resultado e discussão
A aproximação dos valores entre as poses da estrutura cristalográfica do
ligante e dos confôrmeros dos ligantes estudados pode fornecer dados
relevantes para o conhecimento da interação da estrutura com o alvo
biológico (YE, LING, CHEN, 2017). A Figura 2 mostra os resultados do docking
para os receptores AChE e Hormônio Juvenil.
A piperina apresenta 3 subunidades: um anel 1,3-benzodioxola, unido a uma
cadeia de butadieno com função cetona -insaturada e, este ligado ao
fragmento amida (Figura 1).
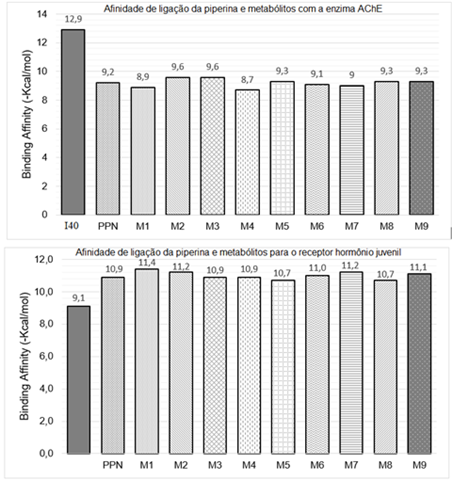
Ao analisar as energias teóricas de afinidade de ligação na a Figura 1,
nota-se que todos os metabólitos obtidos via PreADMET para piperina,
apresentaram menor afinidade de ligação com a enzima AChE de Drosophila
melanogaster (PDB ID 1QON), demonstrando que estes não podem ser
considerados potenciais agentes larvicidas para o mosquito Ae. aegypti.
Por sua vez, a piperina e metabólitos exibiram alta afinidade de ligação
para o receptor hormônio juvenil (PDB ID 5V13). O metabólito M1 (- 11,4
Kcal/mol), exibiu o maior valor de energia de ligação indicando que a
epoxidação próximo ao núcleo benzodioxol da piperina incrementa a afinidade
quando a comparada a outras posições na estrutura. A hidroxilação também
mostrou ótimos valores de afinidade: M2 e M7 (- 11,2 Kcal/mol) e M9 (- 11,1
Kcal/mol), indicando que tanto a introdução do grupo hidroxila em qualquer
parte da cadeia butadieno ou na posição meta do anel piperidínico promove o
aumento da afinidade de ligação. Os resultados mostrados foram superiores a
valores de outros compostos encontrados na literatura incluindo controles de
inibição como JHIII e piriproxifeno (RAMOS et al., 2019).
Conclusões
Os metabólitos da piperina são os prováveis responsáveis pela atividade inseticida do alcaloide. Reações de epoxidação e hidroxilação aumentam a afinidade de ligação entre os ligantes e o sítio enzimático, sendo superiores ao valor de energia de ligação da piperina. A epoxidação próximo ao núcleo benzodioxol da piperina e a introdução dos grupos hidroxila em qualquer parte da cadeia aberta insaturada ou na posição meta da subunidade piperidínica são responsáveis pelo aumento da afinidade de ligação com os receptores hormônio juvenil e acetilcolinesterase.
Agradecimentos
Referências
BIOVIA. Discovery Studio, v.17.2.0. Modeling Environment, Release 2017. Dessault Systèmes, San Diego, CA, USA, 2016.
BLENAU, W.; RADEMACHER, E.; BAUMANN, A. Plant essential oils and formamidines as insecticides/acaricides: what are the molecular targets? Apidologie 2012, 43(3), 334–347, 2012. doi: 10.1007/s13592-011-0108-7.
BOTAS, G. S.; CRUZ, R. A. S.; DE ALMEIDA, F. B.; DUARTE, J. L.; ARAÚJO, R. S.; SOUTO, R. N. P.; FERREIRA, R.; CARVALHO, J. C. T.; SANTOS, M. G.; ROCHA, L.; PEREIRA, V. L. P.; FERNANDES, C. P. Baccharis reticularia DC. and limonene nanoemulsions: Promising larvicidal agents for Aedes aegypti (Diptera: Culicidae) control. Molecules 22, 1990, 2017. doi:10.3390/molecules22111990.
CHEMOFFICE. ChemOffice 3D Ultra 7.0 for Windows. Cambridge, USA: CambridgeSoft Corporation, 2005.
COSTA, G. V.; FERREIRA, E. F. B.; RAMOS, R. S.; SILVA, L. B.; SÁ, E. M. F.; SILVA, A. K. P.; LOBATO, C. M.; SOUTO, R. N. P.; SILVA, C. H. T. P.; FEDERICO, L. B.; ROSA, J. M. C.; SANTOS, C. B. R. Hierarchical virtual screening of potential insectides inhibitors of acetylcholinesterase and juvenile hormone from temephos. Pharmaceuticals 12, 61, 2019. doi:10.3390/ph12020061.
ENGDAHL, C.; KNUTSSON, S.; FREDRIKSSON, S-Å.; LINUSSON, A.; BUCHT, G.; EKSTRÖM, F. Acetylcholinesterases from the disease vectors Aedes aegypti and Anopheles gambiae: Functional characterization and comparisons with vertebrate orthologues. PLoS ONE 10(10), e0138598, 2015. doi:10.1371/journal.pone.0138598.
GIACOPPO, J. O. S., CARREGAL, J. B., JUNIOR, M. C., CUNHA, E. F. F.; RAMALHO, T. C. Towards the understanding of tetrahydroquinolines action in Aedes aegypti: Larvicide or adulticide? Mol. Simulat. 43(2), 121–133, 2017. doi: 10.1080/08927022.2016.1239823.
GORGANI, L.; MOHAMMADI, M.; NAJAFPOUR, G.; NIKZAD, M. Piperine - The bioactive compound of black pepper: From isolation to medicinal formulations. Compr. Ver. Food Sci. F. v. 00, p. 1–17, 2017. doi: 10.1111/1541-4337.12246.
GROOM, C. R.; BRUNO, I. J.; LIGHTFOOT, M. P; WARD, S. C. The Cambridge Structural Database. Acta Cryst. B72, 171–179, 2016. doi: 10.1107/S2052520616003954.
HAMID, P. H.; NINDITYA, V. I.; PRASTOWO, J.; HARYANTO, A.; TAUBERT, A.; HERMOSILLA, C. Current Status of Aedes aegypti insecticide resistance development from Banjarmasin, Kalimantan, Indonesia. BioMed Res. Int., 1–7. 2018. doi:10.1155/2018/1735358.
HYPERCUB INC. HyperChem Program Release 8.0.7 for Windows. Gainsville, FL, 2009.
KROUPOVA et al., 2018 - KROUPOVA, A.; IVASCU, A.; REIMAO-PINTO, M. M.; AMERES, S. L.; JINEK, M. Structural basis for acceptor RNA substrate selectivity of the 3’ terminal uridylyl transferase Tailor. Nucleic Acids Res. 47 (2), 1030–1042, 2018. doi: 10.1093/nar/gky1164.
LINNAEUS, C. Zweyter Theil, enthalt Beschreibungen verschiedener wichtiger naturalien. p. 267-606. In: HASSELQUIST, F. Reise nach Palastina in den Jaren von 1749 bis 1752, 1762. Rostock, Germany, 1762.
MARQUES, A. M; KAPLAN, M. A. C. Active metabolites of the genus Piper against Aedes aegypti: Natural alternative sources for dengue vector control. Univ. Sci. 20(1), 61–82, 2015. doi:10.11144/javeriana.sc20-1.amgp.
PAHO. Pan American Health Organization. Reported cases of dengue fever in the Americas, by country or territory by 2016-2019. (until July 2019), 2019.
RAMOS R. S.; COSTA, J. S.; SILVA, R. C.; COSTA, G. V.; RODRIGUES, A. B. L.; RABELO, É. M.; SOUTO, R. N. P.; TAFT, C. A.; SILVA, C. H. T. P.; ROSA, J. M. C.; SANTOS, C. B. R.; MACÊDO, W. J. C.. Identification of potential inhibitors from pyriproxyfen with insecticidal activity by virtual screening. Pharmaceuticals 12(1); 20, 2019. doi: 10.3390/ph12010020.
RAMOS-CASTAÑEDA J.; SANTOS, F. B., MARTÕÂNEZ-VEGA, R.; ARAUJO, J. M. G.; JOINT, G.; SARTI, E. Dengue in Latin America: Systematic review of molecular epidemiological trends. PLoS Negl. Trop. Dis. 11(1), e0005224, 2017. doi:10.1371/journal.pntd.0005224.
ROIZ D.; WILSON A. L.; SCOTT, T. W.; FONSECA, D. M.; JOURDAIN, F.; MÜLLER, P.; VELAYUDHAN, R.; CORBEL, V. Integrated Aedes management for the control of Aedes-borne diseases. PLoS Negl. Trop. Dis. 12(12), e0006845, 2018. doi: 10.1371/journal.pntd.0006845.
TROTT, O.; OLSON, A. J. Software news and update AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comp. Chem. 31, 455–461, 2010.
VENKATESH; SHASTRI, S. L.; KRISHNA, V.; JAYABASKARAN, C. In Silico analysis of ADME-T properties of amentoflavone. Bioinform. Proteom. Opn. Acc. J. 1(3), 000118, 2017.
YE, X.-Y.; LING, Q.-Z.; CHEN, S.-J. Identification of neprilysin as a potential target of arteannuin using computational drug repositioning. Braz. J. Pharm. Sci. 53(2), e16087, 2017. doi: 10.1590/s2175-97902017000216087.
ZARA, A. L. S. A.; SANTOS,S. M.; FERNANDES-OLIVEIRA, E. S.; CARVALHO, R.G.; COELHO, G. E. Estratégias de controle do Aedes aegypti: Uma revisão. Epidemiol. Serv. Saude 25(2), 391–404, 2016. doi: 10.5123/S1679-49742016000200017.








